Dê sua opinião
Outros Idiomas
English
Español
Français
Interlingua
|
|
ECOPOESE.
Resumo - O modelo da Ecopoese
é uma hipótese sobre a origem da
vida na Terra que propõe que os ciclos
geoquímicos dos elementos biogênicos
(ou ciclos biogeoquímicos) precederam o
aparecimento dos organismos. A existência
de tais ciclos em um mundo prebiológico
implica a presença de fortes doadores (redutores)
e aceptores (oxidantes) de elétrons, reabastecidos
por meio de reações abióticas
no ambiente aquoso primitivo da terra.
A ciclagem de compostos de carbono, nitrogênio
e enxofre ocorreria nos oceanos, um ambiente aquoso
em contato com uma litosfera rica em íons
redutores, tais como o ferro ferroso (divalente)
e sulfeto, e uma atmosfera contendo quantidades
significativas de oxigênio molecular (livre),
gerado pela fotólise atmosférica
do vapor d'água.
As transições redox pertinentes
aos processos vivos seriam assim atributos primordiais
da ecosfera nascente, e determinariam a natureza
dos materiais e dos processos de obtenção
que moldariam gradativamente a evolução
da organismalidade .
Níveis atmosféricos altos de gás
carbônico seriam responsáveis por
algumas propriedades reativas essenciais deste
mundo protobiológico, tais como a possibilidade
de fixação de carbono, e propagação
quiral.
O modelo da Ecopoese (Félix de Sousa,
R.A., 2000, 2006) para a origem da vida se fundamenta
na idéia de que uma atmosfera contendo
uma alta porcentagem de oxigênio molecular,
gerado pela fotólise atmosférica
do vapor d'água, é uma característica
essencial da ecosfera primitiva da Terra.
A Ecopoese oferece uma perspectiva diferente com
relação aos outros modelos de biopoese.
Tradicionalmente, a origem da vida é considerada
equivalente ao surgimento casual de organismos
celulares muito simples, cuja evolução
metabólica iria, daí por diante
guiar a evolução geral do ambiente
(a atmosfera rica em oxigênio seria um resultado
deste processo). Na ecopoese, em contraste, o
ambiente físico desempenha o principal
papel, e não os organismos.
Além de dotar o meio ambiente, desde o
início, com a proteção contínua
de uma camada de ozônio, a presença
precoce do oxigênio livre determina o estabelecimento
de um fluxo de elétrons que, deixando os
componentes redutores da litosfera, cruzam os
mares arqueanos, bombeados pela produção
fotolítica de oxigênio na atmosfera
terrestre. Este fluxo é mediado e em grande
parte canalizado pelas transições
redox dos elementos redox-sensíveis na
hidrosfera.
Estas interações ambientais de grande
escala ocasionam o desenvolvimento de um metabolismo
de base geoquímica em um protoplasma planetário
(o holoplasma) criando as pré-condições
para a evolução gradual da vida
organismal.
|
A ampla diferença de potenciais
eletroquímicos no ambiente primordial causaria
o aparecimento dos ciclos geoquímicos dos elementos
biogênicos. As vias metabólicas primitivas
se originam da interação destes ciclos
e os seus produtos. A geração de ordem
no sistema decorre das transições energeticamente
favoráveis, particularmente na oxidação
da matéria orgânica, e das propriedades
físico-químicas das substâncias
envolvidas. Este protometabolismo planetário
é essencialmente congruente à bioquímica
dos organismos atuais, incluindo a fixação
de carbono e nitrogênio, e a degradação
aeróbica de compostos orgânicos (oxidação
total a CO2).
A evolução biológica, em regra,
procederia pelo controle funcional crescente destas
reações, e não por sua criação.
Um metabolismo nu de base geoquímica evoluiria
assim, de modo congruente, em direção
aos processos enzimáticos modernos.
|
CICLOS GEOQUÍMICOS
DOS ELEMENTOS BIOGÊNICOS NO
PERÍODO ARQUEANO.

|
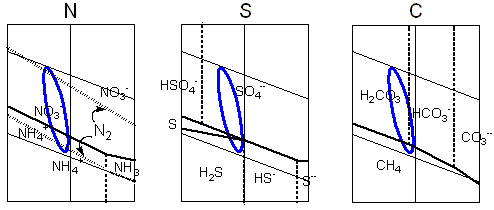
A gama de potenciais eletroquímicos na Terra protobiológica é calculada como variando entre os valores extremos determinados pelo Ferro divalente da litosfera e o oxigênio atmosférico.
Os ciclos geoquímicos primitivos são constituídos pelas espécies químicas possíveis dos elementos biogênicos e suas transições neste intervalo.
A natureza oxidante da atmosfera primitiva decorreria da produção abiótica de oxigênio, por fotólise do vapor d'água atmosférico e perda gravitacional de hidrogênio.
A baixíssima solubilidade do oxigênio nas condições de alta temperatura dos oceanos primitivos e a relativa escassez de terras emersas exigiriam a manutenção de uma elevada concentração de oxigênio na atmosfera, necessária para que o fluxo de elétrons a partir dos compartimentos hidrosférico e litosférico pudessem compensar a perda por fotólise.
A maior parte dos gases redutores originados pela atividade vulcânica seria liberada diretamente na hidrosfera ou teria curta residência na atmosfera. Pequenas quantidades de metano ou monóxido de carbono, menos solúveis, poderiam entretanto coexistir em equilíbrio com a atmosfera oxidante.
As áreas de estabilidade para as espécies químicas derivadas dos elementos biogênicos estão representadas nos diagramas de Pourbaix. A linha azul liga os valores extremos de pH e de potencial eletroquímico calculados para o meio hipercarbônico. Pretende ilustrar, de modo aproximado, as transições químicas possíveis nas condições protobiológicas propostas.
No caso do enxôfre os ciclos seriam complicados pela interferência com o ferro divalente. O diagrama de Pourbaix para o carbono inclui somente os dois extremos, a espécie mais reduzida - o metano, e a mais oxidada, o dióxido de carbono (e seus derivados aquosos, o ácido carbônico, e os íons bicarbonato e carbonato), entre um grande número de compostos possíveis exibindo estados de redução intermediários. |
|
| |

|
|
As propriedades carboxilantes e condensantes
emprestadas ao médio aquoso (apelidado "hipercarbônico")
pela presença de uma densa atmosfera de CO2 são particularmente importantes, pois permitem
a fixação de carbono na presença
do poder redutor, e a propagação quiral.
|
UM MUNDO HIPERCARBÔNICO.
| |
A
expressão "meio hipercarbônico"
descreve as propriedades reativas do ambiente
oceânico da Terra primitiva sob a composição
atmosférica postulada no modelo da ecopoese.
Devido à grande quantidade de CO2
dissolvido, especies químicas tais como
o ácido carbônico, o íon bicarbonato
e íons divalentes (principalmente o alcalino-terroso
magnésio) estão presentes em concentrações
bem mais altas que nos oceanos hodiernos, emprestando
propriedades carboxilantes capazes de afetar grupamentos
amino e carbânions Sob o equilíbrio carboxilação-descarboxilação assim determinado, vários compostos orgânicos formariam séries de "análogos hipercarbônicos" estreitamente assemelhados, cujos membros diferem unicamente quanto à quantidade de CO2 adicionada às suas moléculas.

O
meio altamente polar com baixa atividade de agua
(Aw) governa o equilíbrio
de hidratação-desidratação.
Adicionalmente, um equilíbrio redução-oxidação
é parte do ambiente oceânico como um
todo, posicionado entre uma litosfera que fornece
íons redutores (sobretudo ferro e enxofre
divalentes) e a atmosfera óxica. Os fatores
que determinam o equilíbrio de carboxilação-descarboxilação
e o equilíbrio hidratação-desidratação
são designados fatores permanentes e estão
ligados a condições que somente flutuam
muito levemente. Por outro lado o equilíbrio
oxidação-redução é
governado pelos fatores eventuais, que variam grandemente,
entre os extremos redox do ambiente protobiológico.
A química do meio hipercarbônico é central ao modelo da ecopoese pois concilia, tal como ditado pelo princípio da congruência, a relevância biológica das reações de carboxilação e descarboxilação com as evidências geológicas de uma paleoatmosfera pesada de CO2. |
|
Como o dióxido
de carbono é extremamente estável
com respeito à reação de redução
em soluções aquosas, a fixação
de carbono dependeria da carboxilação
de compostos de carbono preexistentes. Os ciclos
protobiológicos do carbono teriam, no sentido
anabólico, reações de carboxilação
intercaladas com etapas de redução.
Inversamente, os ciclos catabólicos alternariam
descarboxilações e etapas de oxidação,
como nos ciclos metabólicos modernos. Os
possíveis análogos não-enzimáticos
destas reações no meio hipercarbônico
são discutidos com algum detalhe no livro,
bem como os mecanismos que fariam com que o equilíbrio
redução-oxidação na
ecosfera protobiológica estivesse firmemente
ligado aos processos de carboxilação
e descarboxilação.
|

A fixação de carbono
dependeria também de uma contribuição,
ao menos modesta (aporte primário), de hidrocarbonetos
formados pela decomposição de carbetos
procedentes da litosfera. O acetileno, em particular
poderia ser convertido diretamente, por hidratação
e carboxilação, nos ácidos pirúvico
e oxaloacético, constituintes centrais das vias
bioquímicas atuais. A participação
destes compostos reforça a idéia de que
o ciclo de Krebs bidirecional seria a característica
central dos processos de circulação
de carbono na ecosfera primitiva.
A disponibilidade de poder redutor para os processos
anabólicos (derivando principalmente do ferro
divalente e dos sulfetos litosféricos), comparada
à do oxigênio atmosférico apenas
ligeiramente solúvel, favoreceria a acumulação
de compostos orgânicos na hidrosfera.
O questionamento a respeito da natureza destes compostos
e de seus processos de formação pode ser
auxiliada pela presunção de uma evolução
congruente.
|
O PRINCÍPIO DA
CONGRUÊNCIA.
| |
O princípio da congruência é uma ferramenta heurística destinada a guiar a investigação da história dos sistemas vivos. Enuncia simplesmente que a vida moderna é uma ferramenta necessária e utilíssima no estudo dos estados anteriores da vida. A congruência pode ser encarada como um complemento à teoria da evolução pois representa a busca de evidências de ancestralidade comum num nível molecular.
Embora as similaridades bioquímicas entre os organismos existentes sejam geralmente considerados uma prova inequívoca de sua evolução comum, os modelos clássicos de origem da vida propoem um mundo biológico anterior, ou "prebiótico", onde os compostos orgânicos teriam se acumulado por meio de processos estranhos à bioquímica hodierna.
As primeiras proposições teóricas com relação à pesquisa da natureza evolutiva do metabolismo foram estabelecidas por Granick nos anos 40 e mais recentemente aplicadas, notadamente por De Duve e por Morowitz ao campo da origem da vida. O modelo da Ecopoese estende ainda mais além os domínios da congruência, da evolução dos organismos à da ecosfera como um todo.
Esta concepção implica que os ciclos
geoquímicos dos elementos biogênicos
foram sempre essencialmente os mesmos, mas também
que os íngremes gradientes de energia que
aprovisionam os processos vitais não foram
criados pelos organismos. Ao contrário, é
exatamente a existência de processos geradores
de energia no ambiente primordial, especialmente
a degradação aeróbica da matéria
orgânica (oxidação total a CO2),
que teria permitido a criação da ordem
inerente à vida organismal.
A utilização do termo "protobiológico", implicando a vida primitiva, no lugar de "prebiótico" (antes da vida) enfatiza o tipo de continuidade que caracteriza o princípio da congruência e é consistentemente seguida no modelo da ecopoese.. |
|
A congruência pressupõe analogia química
entre estas duas fases da evolução bioquímica, e é demonstrado o paralelismo
entre a ação dos fatores ambientais protobiológicos
e das coenzimas.
|
PROCESSOS CONGRUENTES
DO MUNDO HIPERCARBÔNICO.
| |
Fatores reativos ambientais e reativos metabólicos correspondentes.
1 - Permanentes
Fator ambiental (e reação contrária) |
Reativos metabólicos correspondentes |
Origem |
+ H2O
- H2O
|
(espontâneo)
ATP, GTP, etc. |
Hidroatmosfera
|
+ CO2
- CO2
|
Biotina
(espontâneo) |
Hidroatmosfera
|
2 - Eventuais primários.
[H]
[O]
|
TH - Transportadores Reduzidos
(NADH, FADH2, etc.)
T - Transportadores Oxidados
(NAD+, FAD, etc.) |
Poder Redutor Litosférico
Oxigênio Atmosférico |
+
H2S
- H2S
(+H2O)
|
CoenzimaA-SH
(espontâneo) |
Hidroatmosfera (Ciclo do S) |
+
NH3 (NH2COOH)
- NH3
(+H2O)
|
Compostos aminados
(espontâneo) |
Hidroatmosfera (Ciclo do N) |
+
H2PO4-
-H2PO4-
(+H2O)
|
ATP, GTP, etc.
(espontâneo) |
Hidroatmosfera |
3 - Eventuais secundários.
H2S2 |
Ácido Lipóico |
H2S
/ [O] |
NH2COOH
/ [H]
(NH2COSH / [H])
Nucleófilo de Nitrogênio |
Piridoxamina |
NH2COOH
/ [H] |
CN-
Nucleófilo de Carbono |
Tiamina |
Cianeto
ou derivados do ácido fórmico
(Um-carbono) |
|
|
As coenzimas representam um primeiro degrau no
sentido da incorporação do metabolismo
nu primordial em unidades portadoras de atividade
catalítica. A probabilidade de que esta
transição tenha ocorrido principalmente
durante o "mundo do ARN", onde as funções
metabólicas eram realizadas por ribozimas,
é documentada pelo fato de que a maioria
destas coenzimas é estruturalmente relacionada
com os nucleosídeos que contêm ribose.
Os tópicos com asterisco (*) exigem um
tratamento mais profundo, só encontrado
no livro.
A propagação quiral (polimerização
enantioseletiva) garantiria a síntese de
polipeptídeos homoquirais devido à
formação de centros proquirais nos
derivados hipercarbônicos dos materiais
peptídicos.
|
PROPAGAÇÃO QUIRAL NO MEIO HIPERCARBÔNICO.
| |

O mecanismo de propagação
quiral aqui apresentado para aminoácidos
racêmicos é uma versão minimamente
desnudada das formas de equilíbrio possíveis
no meio hipercarbônico. Em conformidade, a
N-carboxilação e a protonação
da carboxila da cadeia principal são essenciais,
mas as verdadeiras espécies reativas podem
demandar a adição de CO2
ou H2S ao material peptídico,
com a utilização dos aminodiácidos
ou tioácidos correspondentes. Excetuando
o fato de que os organismos modernos utilizam aminoácidos
opticamente ativos, o mecanismo é em todos
os aspectos completamente congruente com a bioquímica
atual.
No meio hipercarbônico, os aminoácidos estão destituídos de características zwitteriônicas. O ataque nucleofílico do grupo amino carboxilado sobre a carboxila protonada procede facilmente, gerando um aduto que contém um carbânion estabilizado por ressonância. A recombinação assimétrica com o próton é orientada pelo aminoácido adicionado no ciclo anterior. Como a iniciação pode ser feita por acilação com um ácido carboxílico aquiral, o primeiro aminoácido da cadeia poderá ser qualquer um dos dois enantiômeros, mas determinará a orientação quiral de todos os subseqüentes, produzindo polipeptídeos homoquirais.
Outro aspecto importante é que a acilação do grupamento amino do fragmento de aminoácido provoca a elevação do pka do grupamento carboxila aumentando a probabilidade de que esteja protonado e favorecendo assim o alongamento de cadeias já em formação em detrimento do ataque a monômeros.
A teoria mecanística da origem da quiralidade no mundo biológico é parte integrante do modelo da ecopoese. |
|
As características organismais de vida só
serão adquiridas gradualmente, e o advento das
células é bastante tardio neste modelo.
|
EVOLUÇÃO
DA ORGANISMALIDADE: OS HÁBITOS PROTOBIOLÓGICOS.
| |
MATERIAIS PROTOBIOLÓGICOS E
O HÁBITO FLOCULAR

A agregação
inicial de material orgânico (chamada "hábito
flocular"), contrariando a tendência
entrópica, é atribuída a uma
combinação de fatores. A baixa atividade
de água (Aw)
A agregação
inicial de material orgânico (chamada "hábito
flocular"), contrariando a tendência
entrópica, é atribuída a uma
combinação de fatores:
1) A baixa atividade
de água (Aw) do meio hipercarbônico.
2) A coalescência
dos compostos hidrofóbicos (minimizando a
energia devida à separação
de cargas no meio altamente polar).
3) A cinética
de formação de tioácidos e
a sua extração parcial para a fase
lipídica.
4) A estabilização
da micela hidrofóbica pela formação
de compostos anfifílicos em sua superfície.
5) A fácil
formação de ligações
peptídicas devida à destruição
do caráter zwitteriônico dos aminoácidos
pela N-carboxilação.
6) A adesão
da fração peptídica aos compostos
anfifílicos superficiais da micela devida
à afinidade de cargas, com a provável
participação de cátions polivalentes.
7) A adsorção
de compostos orgânicos de baixo peso molecular
à fração peptídica.
Todas estas
propriedades contribuiriam conjuntamente na estabilização
de agregados coloidais numa solução
diluída. Coadunam-se inclusive com a visão
da Terra primitiva como um "mundo de águas",
destituído de superfícies de evaporação
significativas.
A solubilidade preferencial de oxigênio na
fase lipídica favorece os processos de oxidação
próximos à superfície da micela.
Um processo bastante simples, a fosforilação
homogênea - análogo à fosforilação
a nível do substrato, demandando a oxidação
do sulfeto de hidrogênio a dissulfeto de hidrogênio
(análogos do ácido lipóico),
causaria a oxidação de a-cetoácidos
a tioácidos, permitindo a síntese
de compostos de 'alta-energia'. Aminoácidos
contendo enxofre (grupamentos SH) na fração
peptídica poderiam ligar-se ao ferro divalente
de origem litosférica. Estes agregados ferro-enxofre
primitivos, ancestrais das modernas ferredoxinas,
são os responsáveis pela concentração
do poder redutor, concentrando os potenciais redox
nos agregados coloidais primitivos, que teriam assim
tanto polos redutores como oxidantes.

Uma interação inicialmente frouxa
e genérica entre os polipeptídeos
e os polímeros fosfodiéster (PFDE
- ou RNA primitivo) aumentaria em especificidade
dando origem à tradução, e
ao controle estrutural e funcional que caracteriza
o "hábito reticular".
O HÁBITO RETICULAR.

Embora a fosforilação
heterogênea ("respiração")
exija barreiras físicas, como membranas,
seu desenvolvimento poderia ter ocorrido antes do
advento do "hábito celular", pois
os prótons que resultam da oxidação
de transportadores de elétrons lipossolúveis
seriam segregados em vacúolos protônicos
(ou ácidos), criando assim um potencial entre
este vacúolo e o compartimento externo (ou
metabólico), o que permitiria o desenvolvimento
de um método mais eficiente para a síntese
de compostos de alta-energia.
CELULARIDADE E RESPIRAÇÃO
CELULAR.

No modelo da Ecopoese a celularidade não
surge como um método para proteger os compostos
orgânicos da diluição, mas para
promover sua oxidação e tirar proveito
da mesma. Esta proposição de uma aurora
aeróbica para a celularidade é fundamentada
nas propriedades funcionais energéticas das
células, mais do que em sua aparência
física.
HÁBITOS CELULARES.

O fechamento gradual do recesso metabólico conduziria
ao hábito celular (onde o vacúolo protônico
não é mais necessário). |
|
O LIVRO.
 |
Clique aqui para baixar ECOPOESE - A criação
da ecosfera, 2ª Ed., 2006; - PDF Grátis
(1,7 Mb, 345 páginas).
Ou deixe uma mensagem para
comprar a versão impressa.
Este documento é protegido
por direito autoral. Entretanto, quaisquer partes
da obra podem ser livremente reproduzidas ou traduzidas
para propósitos não-comerciais,
desde que a fonte seja claramente referenciada.
Este livro deve ser referenciado
do seguinte modo:
Félix de Sousa, Raul A., (2006). Ecopoese
- A criação da ecosfera, 2ª
Ed., Rio de Janeiro. |
UM ARTIGO.
Uma versão condensada viXra
em inglês pode ser baixada aqui
( PDF Gratuito 526 kb, 39 páginas ).
Para referenciá-la:
Félix de Sousa, Raul A. (2010). The
Ecopoesis Model: Did Free Oxygen Fuel the Origin of Life?
viXra: 1010.0001
|
 ECOPOESE
ECOPOESE
