Su opinión
Otros Idiomas
English
Français
Interlingua
Português
|
|
ECOPOIESIS.
Resumen - El modelo de la
Ecopoiesis es una hipótesis sobre el origen
de vida en la Tierra que propone que los ciclos
geoquímicos de los elementos biogénicos
(o ciclos biogeoquímicos) precedieran al
aparecimiento de los organismos. La existencia
de tales ciclos en un mundo prebiológico
implica la presencia de fuertes donadores (reductores)
y aceptadores (oxidantes) de electrones, reabastecidos
por reacciones abióticas en el ambiente
acuoso primitivo de la Tierra.
El ciclaje de compuestos de carbono, nitrógeno
y azufre tendría lugar en los océanos,
un ambiente acuoso en contacto con una litósfera
rica en iones reductores como el hierro ferroso
(divalente) y sulfuro, y una atmósfera
conteniendo cantidades sustanciales de oxígeno
molecular (libre), generado por la fotólisis
atmosférica del vapor de agua.
Las transiciones redox pertinentes a los procesos
vivientes serían así atributos primordiales
de la ecósfera naciente, determinando la
naturaleza de los materiales y procesos de obtención
de energía que amoldarían la evolución
gradual de la organismalidad.
Niveles atmosféricos altos de anhídrido
carbónico responderían por algunas
propiedades reactivas esenciales de este mundo
protobiológico, como la posibilidad de
fijación del carbono, y la propagación
quiral.
El modelo de la Ecopoiesis (Félix de Sousa,
R.A., 2000, 2006) para el origen de la vida es
basado en la idea que una atmósfera conteniendo
un porcentaje alto de oxígeno molecular,
generado por la fotólisis atmosférica
del vapor de agua, es un rasgo esencial de la
ecosfera primitiva de la Tierra.
La Ecopoiesis ofrece una perspectiva distincta
de los otros modelos de biopoesis. Tradicionalmente,
el origen de la vida es tenido como equivalente
al aparecimiento afortunado de organismos celulares
muy simples, cuya evolución metabólica
iría desde entonces conducir la evolución
general del ambiente (la atmósfera rica
en oxígeno seria resultado de este proceso).
En la ecopoiesis, en contraste, el ambiente físico
juega el papel principal, y no los organismos.
Además de dotar el medio ambiente, desde
el inicio, con la protección continua de
una capa de ozono, la presencia temprana de oxígeno
libre determina el establecimiento de un flujo
de electrones que, dejando los componentes reductores
de la litosfera, cruzan los mares arqueanos, bombeados
por la producción fotolítica de
oxígeno en la atmósfera de la Tierra.
Este flujo es mediado y en grande parte canalizado
por las transiciones redox de los elementos redox-sensibles
en la hidrosfera.
Esas interacciones medioambientales de gran amplitud
causan el desarrollo de un metabolismo de base
geoquímica en un protoplasma planetario
(el holoplasma) creando las pre-condiciones para
la evolución gradual de la vida organismal.
|
La amplia diferencia
de potenciales electroquímicos en el ambiente
primordial causaría el aparecimiento de los
ciclos geoquímicos de los elementos biogénicos.
Las vías metabólicas primitivas se
originan de la interacción de estos ciclos
y sus productos. El aumento de orden en el sistema
sobreviene de las transiciones enérgicamente
favorables, particularmente en la oxidación
de materia orgánica, y de las propiedades
físico-químicas de los compuestos
participantes. Este protometabolismo planetario
es esencialmente congruente con la bioquímica
de hoy, incluyendo la fijación de carbono
y de nitrógeno, y la degradación aeróbica
de los compuestos orgánicos (oxidación
total a CO2).
La evolución biológica, como regla,
procedería a través del control funcional
creciente de estas reacciones, y no por su creación.
Un metabolismo desnudo de base geoquímica
evolucionaría así congruentemente
hacia nuestros procesos enzimáticos modernos.
|
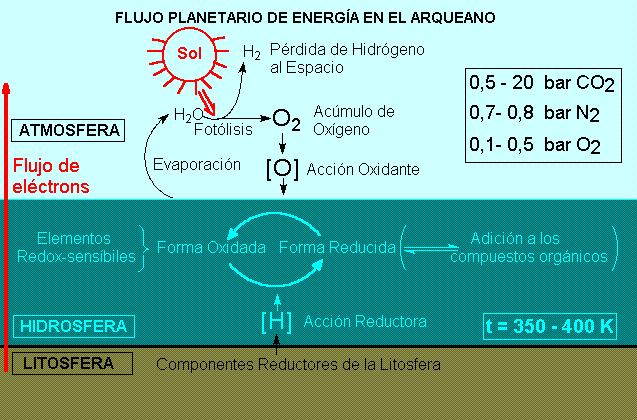
CICLOS GEOQUÍMICOS
DE LOS ELEMENTOS BIOGÉNICOS EN EL
PERIODO ARQUEANO.
|
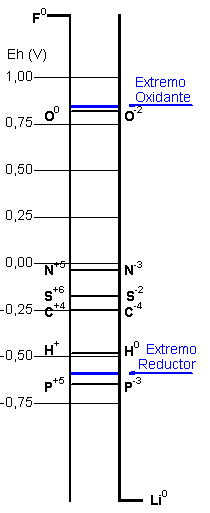
El rango de potenciales electroquímicos en la Tierra protobiológica es calculado como variando entre los valores del hierro bivalente litosférico a los del oxígeno atmosférico.
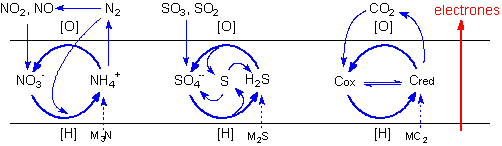
Los primeros ciclos geoquímicos son constituidos de las especies químicas posibles para los elementos biogénicos y de sus transiciones dentro de este intervalo.
La naturaleza oxidante de la atmósfera primitiva sería determinada por la producción abiótica de oxígeno, originado de la fotólisis del vapor de agua atmosférico seguido por la pérdida gravitacional de hidrógeno.
La muy baja solubilidad del oxígeno en los océanos primordiales calientes y la escasez de tierras emersas requerirían el mantenimiento de una alta concentración del oxígeno en la atmósfera, para que el flujo de electrones de los compartimientos hidrosférico y litosférico pudiera compensar la pérdida con la fotólisis.
La mayoría de los gases reductores originados de actividad volcánica serían liberados directamente a la hidrosfera o tendrían residencia corta en la atmósfera. Pequeñas cantidades de metano o de monóxido de carbono, menos solubles podrían, sin embargo, coexistir en equilibrio con la atmósfera oxidante.
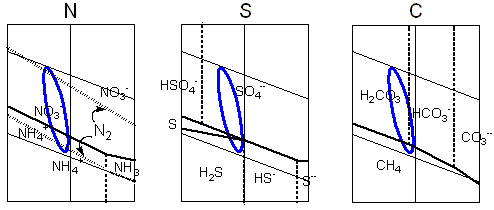
Las áreas de estabilidad para las espécies químicas derivadas de los elementos biogénicos son trazadas en los diagramas de Pourbaix. La línea azul liga los valores extremos de pH y de potencial electroquímico calculados para el medio hipercarbónico. Pretende representar, en una visión aproximada, las transiciones químicas posibles en las condiciones protobiológicas propuestas. En el caso del azufre los ciclos serían complicados por las interacciones con el hierro bivalente. El diagrama de Pourbaix para el carbón incluye solo los dos extremos, la especie más reducida - el metano, y la más oxidada, el bióxido de carbono (y sus derivados acuosos, el ácido carbónico, el ion bicarbonato y el ion carbonato), entre múltiplos compuestos posibles que exhiben estados intermedios de reducción.
|
|
| |
|
|
Las propiedades
carboxilantes y condensantes prestadas al medio
acuoso (dicho "hipercarbónico")
por la presencia de una pesada atmósfera
de CO2 es particularmente
importante, puesto que permiten la fijación
de carbono en presencia del poder reductor, y
la propagación quiral.
|
UN MUNDO HIPERCARBÓNICO.
| |
La
expresión “medio hIpercarbónico”
describe las características reactivas
del ambiente oceánico de la tierra temprana
bajo la composición atmosférica
postulada en el modelo de la ecopoiesis. Debido
a la alta cantidad de CO2
disuelto, las especies químicas tales como
el ácido carbónico, el bicarbonato
y los iones bivalentes (principalmente el alcalinotérreo magnesio) están presentes
en concentraciones mucho más altas que
en el océano de hoy, impartiendo características
carboxilantes que pueden afectar los grupos aminados
y los carbaniones. Bajo el equilibrio carboxilación-descarboxilación
así determinado, vários compuestos
orgánicos formarían series de "análogos
hipercarbónicos" cuyos miembros estrechamente
relacionados difieren unicamente cuanto a la cantidad
de CO2 añadida a
sus moléculas..
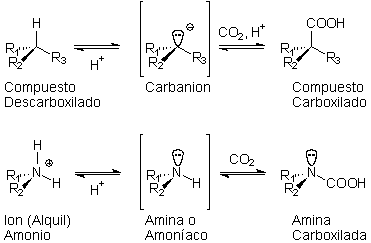
El medio altamente polar con una actividad de agua baja (Aw) gobierna el equilibrio de hidración-deshidratación. Además, un equilibrio de la reducción-oxidación es parte del ambiente oceánico en su totalidad, ubicado entre una litosfera que provee iones reductores (principalmente hierro y sulfuro bivalentes) y la atmósfera óxica. Los factores que determinan el equilibrio de carboxilación-descarboxilación y el equilibrio de hidración-deshidratación se llaman factores permanentes y están vinculados a condiciones que fluctúan muy levemente, mientras que el equilibrio de la reducción-oxidación es gobernado por los factores eventuales, que varían extensamente, entre los extremos redox del ambiente protobiológico.
La química del medio hipercarbónico es central al modelo de la ecopoiesis porque reconcilia, según lo dictado por el principio de la congruencia, la importancia biológica de las reacciones de carboxilación y de descarboxilación con las evidencias geológicas y astronómicas de una pesada paleoatmosfera de CO2. |
|
Como el anhídrido
carbónico es muy estable a la reacción
de reducción en las soluciones acuosas, la
fijación del carbono dependería de
la carboxilación de compuestos de carbono
pre-existentes. Los ciclos protobiológicos
del carbono correrían en el sentido anabólico
intercalando reacciones de carboxilación
de los compuestos orgánicos con etapas de
reducción. Recíprocamente, los ciclos
catabólicos alternarían pasos de descarboxilación
y de oxidación, como en los ciclos metabólicos
modernos. Se discuten las posibilidades de análogos
no-enzimáticos de tales reacciones en el
medio hipercarbónico a un poco de detalle
en el libro, así como los mecanismos a través
de que se ligarían firmemente el equilibrio
de reducción-oxidación en el ecosfera
protobiológica a los procesos de carboxilación
y descarboxilación.
|
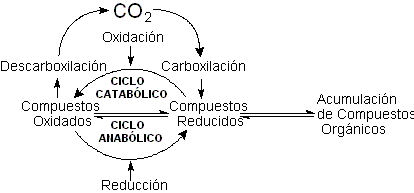
La fijación
del carbono requeriría un aporte por lo menos
modesto (aporte primario) de hidrocarburos formados
por la descomposición de carburos procedentes
de la litosfera, sobre todo el acetileno, que podría
convertirse directamente a través de la hidratación
y carboxilación, a los ácidos pirúvico
y oxaloacético, constituyentes centrales
de las vías bioquímicas actuales.
La participación de estos compuestos refuerza
la idea de que el ciclo de Krebs bidireccional seria
el rasgo principal de la circulación de carbono
en la ecosfera primitiva.
La disponibilidad de poder reductor para los procesos
anabólicos (derivando principalmente del
hierro divalente y de los sulfuros litosféricos),
comparada a la del oxígeno atmosférico
apenas soluble, favorecería la acumulación
de compuestos orgánicos en la hidrosfera.
El questionamiento con respecto a la naturaleza de esos compuestos y de sus procesos
de formación puede ser ayudada por la
suposición de una evolución congruente.
|
EL PRINCIPIO DE LA
CONGRUENCIA.
| |
El
principio de la congruencia es una herramienta heurística
destinada a orientar la investigación de
la historia de sistemas vivos. Enuncía simplemente
que la vida moderna es una referencia altamente
útil y necesaria en el estudio de las etapas
anteriores de la vida. La congruencia se puede considerar
como complemento a la teoría evolutiva puesto
que representa la búsqueda de evidencias
de la ancestralidad común en un nivel bioquímico.
Aunque las semejanzas bioquímicas entre los
organismos existentes sean generalmente considerados una prueba inequívoca
de su evolución común, los modelos
clásicos para el origen de la vida proponen
un mundo anterior, “prebiótico”,
donde los compuestos orgánicos habrían
acumulado en procesos extranos a la bioquímica
de hoy.
Las primeras proposiciones teóricas con respecto
a la investigación de la naturaleza evolutiva
del metabolismo fueron presentadas por Granick en
los años 40, y aplicadas más adelante,
principalmente por De Duve y por Morowitz, al campo
del origen de la vida. El modelo de la Ecopoiesis
amplía más allá el dominio
de la congruencia, de la evolución de organismos
a la de la ecosfera en su totalidad.
Este concepto implica que los ciclos geoquímicos
de elementos biogénicos han sido siempre
esencialmente iguales, pero también que los
íngremes gradientes de energía que
aprovisionan los procesos vitales no han sido creados
por los organismos. En contrario, es la precisamente
la existencia de procesos generadores de energía
en el ambiente primordial, especialmente la degradación
aeróbica de la materia orgánica (oxidación
total al CO2), que habría sido esencial para
la creación de la orden inherente a la vida
organismal.
La utilización del término "protobiológico"
implicando la vida primitiva en lugar de "prebiótico"
(antes de la vida) enfatiza el tipo de continuidad
que caracteriza el principio de la congruencia y
es consistentemente seguida en el modelo de la ecopoiesis. |
|
La congruencia
presupone la analogía química entre
estas dos fases de la evolución bioquímica,
y la acción de los factores protobiológicos
del medio ambiente se apresenta en paralelo con la
de las coenzimas.
|
PROCESOS CONGRUENTES
DEL MUNDO HIPERCARBÓNICO.
| |
Factores
reactivos ambientales y reactivos metabólicos
correspondientes.
1 - Permanentes
Factor
ambiental (y reacción contrária) |
Reactivos
metabólicos correspondientes |
Origen |
+ H2O
- H2O
|
(espontáneo)
ATP, GTP, etc. |
Hidroatmosfera
|
+ CO2
- CO2
|
Biotina
(espontáneo) |
Hidroatmosfera
|
2 - Eventuales
primarios.
[H]
[O]
|
TH
- Transportadores Reducidos
(NADH, FADH2, etc.)
T - Transportadores
Oxidados
(NAD+, FAD, etc.) |
Poder
Reductor Litosférico
Oxígeno
Atmosférico |
+
H2S
- H2S
(+H2O)
|
CoenzimaA-SH
(espontáneo) |
Hidroatmosfera
(Ciclo del S) |
+
NH3 (NH2COOH)
- NH3
(+H2O)
|
Compuestos
aminados
(espontáneo) |
Hidroatmosfera
(Ciclo del N) |
+
H2PO4-
-H2PO4-
(+H2O)
|
ATP,
GTP, etc.
(espontáneo) |
Hidroatmosfera |
3 - Eventuales
secundarios.
H2S2 |
Ácido
Lipóico |
H2S
/ [O] |
NH2COOH
/ [H]
(NH2COSH / [H])
Nucleófilo de Nitrógeno |
Piridoxamina |
NH2COOH
/ [H] |
CN-
Nucleófilo de Carbono |
Tiamina |
Cianuro
o derivados del ácido fórmico
(Un-carbono) |
|
|
Las coenzimas representan un primer peldaño
en la direccíon de la incorporación
del metabolismo nudo primordial en unidades portadoras
de actividad catalítica. La probabilidad
que esa transición haya ocurrido principalmente
durante el "mundo del ARN", donde las
funcciones metabólicas eran realizadas
por ribozimas, es documentada por el facto que
la mayor parte de esas coenzimas es estructuralmente
relacionada con los nucleósidos que contienen
ribosa.
La propagación
quiral (polimerización enantioselectiva)
aseguraría la síntesis de polipéptidos
homoquirales a través de la formación
de centros proquirales en los derivados hipercarbónicos
de los materiales peptídicos.
|
PROPAGACIÓN QUIRAL EN EL MEDIO HIPERCARBÓNICO.
| |
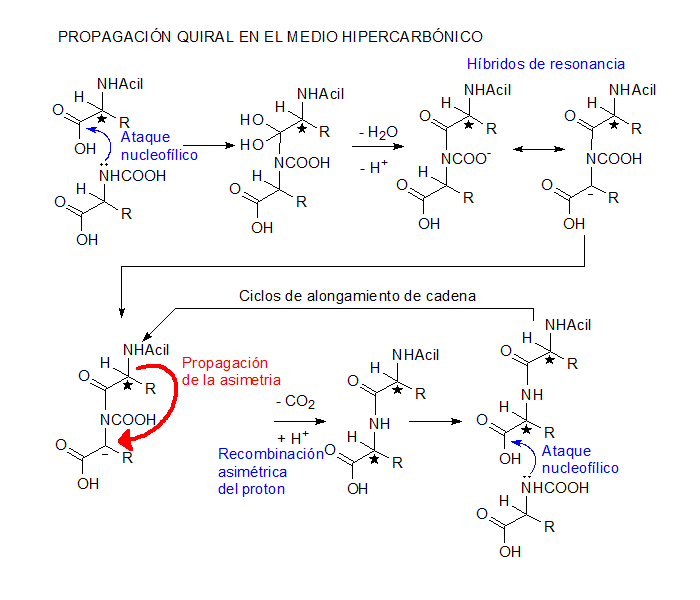
El mecanismo de propagación quiral aqui presentado para aminoácidos racémicos es una version minimamente desnudada de las formas de equilíbrio posíbles en el medio hipercarbónico. En conformidad, la N-carboxilación y la protonación de la carboxila de la cadena principal son esenciales, pero las verdaderas espécies reactivas pueden demandar la adición de CO2 o de H2S al material peptídico, con la utilización de los aminodiácidos o tioácidos correspondientes. Excetuado el facto de que los organismos modernos utilizam aminoácidos opticamente activos, el mecanismo es en todos los aspectos completamente congruente con la bioquímica actual.
En el medio hipercarbónico, los aminoácidos están despojados de características zwitteriónicas. El ataque nucleofílico del grupo amino carboxilado sobre la carboxila protonada procede facilmente, generando un aducto que contiene un carbanion estabilizado por resonancia. La recombinación asimétrica con el proton es orientada por el aminoácido adicionado en el ciclo anterior. Como la iniciación puede ser hecha por acilación con un ácido carboxílico aquiral, el primer aminoácido de la cadena podrá ser qualquiera de los dos enantiómeros, pero determinará la orientación quiral de todos los subsecuentes, producindo polipéptidos homoquirales.
Otro aspecto importante es que la acilación del grupo amino del fragmento de aminoácido provoca la elevación del pka del grupo carboxila aumentando la probabilidad de que esté protonado y favoreciendo asi el alargamiento de cadenas ya en proceso de formación al coste del ataque a monómeros.
La teoría mecanística del origen de la quiralidad en el mundo biológico es una parte integrante del modelo de la ecopoiesis. |
|
Las características
organismales de la vida sólo se adquieren
gradualmente, y el adviento de las células
es bastante tardío en este modelo.
|
EVOLUCIÓN
DE LA ORGANISMALIDAD: LOS HÁBITOS PROTOBIOLÓGICOS.
| |
MATERIALES PROTOBIOLÓGICOS
Y EL HÁBITO FLOCULAR.

La agregación
inicial de la materia orgánica (llamada "hábito
flocular"), contrariando la tendencia entrópica,
se atribuye a una combinación de factores:
1) La baja actividad de agua (Aw)
del medio hipercarbónico.
2) La coalescencia
de los compuestos hidrófobos (minimizando
la energía debido a la separación
de cargas en el medio muy polar).
3) La cinética
de formación de tioácidos y su extracción
parcial en la fase lipídica.
4) La estabilización
de la micela hidrofóbica a través
de la formación de compuestos anfifílicos
en la superficie.
5) La formación
fácil de enlaces peptídicos debido
a la destrucción del carácter zwitteriónico
de los aminoácidos a través de la
N-carboxilación.
6) La adhesión
de la fracción peptídica a los compuestos
anfifílicos superficiales de la micela devida
a la afinidad de cargas, con la probable participación
de cationes polivalentes.
7) A adsorpción
de compuestos orgánicos de bajo peso molecular
a la fracción peptídica.
Todas estas propiedades
contribuirían con la estabilización
de los agregados coloidales en solución diluida.
Se coadunan incluso con la visión de la Tierra
primitiva como un "mundo de aguas", desprovisto
de superficies evaporativas significantes.
La solubilidad preferencial de oxígeno en
la fase lipídica favorece la oxidación
junto a la superficie de la micela. Un proceso bastante
simple, la fosforilación homogénea
- análogo a la fosforilación al nivel
del substrato, requiriendo la oxidación del
sulfuro de hidrógeno al disulfuro de hidrógeno
(análogos del ácido lipóico),
causaría la oxidación de los a-cetoácidos
a los tioácidos, permitiendo la síntesis
de los compuestos de 'alta energía'.
Aminoácidos conteniendo azufre (grupamentos
SH) en la fracción peptídica poderian
ligarse al hierro divalente de origen litosférico.
Estes agregados hierro-azufre primitivos, ancestrales
de las modernas ferredoxinas, son los responsables
por la concentración del poder reductor,
concentrando los potenciales redox en los agregados
coloidales primitivos, que terian así tanto
polos reductores como oxidantes.
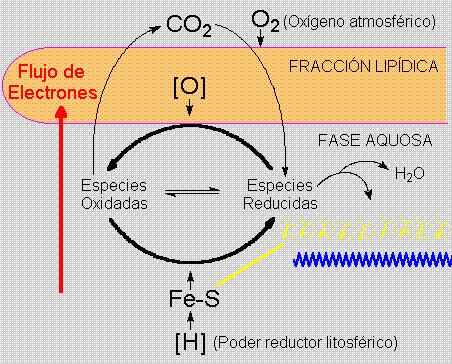
Una interacción inicialmente suelta y genérica
entre los polipéptidos y los polímeros
fosfodiester (PFDE - o ARN primitivo) aumentaría
en especificidad dando lugar a la traducción,
y al control estructural y funcional que caracteriza
el "hábito reticular".
EL HÁBITO RETICULAR.
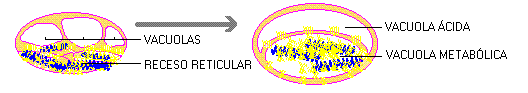
Aunque la fosforilación
heterogénea ("respiración")
requiera barreras físicas, como las membranas,
podría desarrollarse antes del advenimiento
del "hábito celular", ya que los
protones que son el resultado de la oxidación
de los transportadores de electróns solubles
en lípidos serían segregados en la
vacuola protónica (o ácida), creando
así un potencial entre esta vacuola y el
compartimiento externo (o metabólico), lo
que permitiría el desarrollo de un método
más eficaz de síntesis de compuestos
de 'alta energía'.
CELULARIDAD Y RESPIRACIÓN CELULAR.
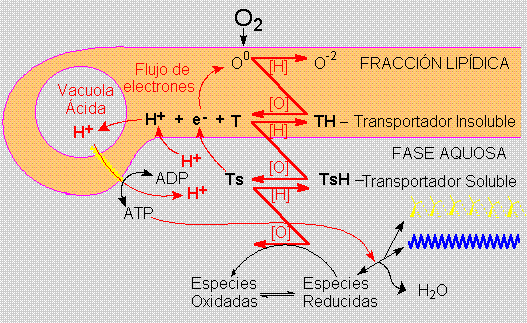
En el modelo de
la Ecopoiesis la celularidad no surge como um método
para proteger los compuestos orgánicos, sino
que para promover y aprovechar de su oxidación.
Esta proposición de una aurora aeróbica
para la celularidad está basada en las propiedades
energéticas funccionales de las células,
más que en su aparéncia física.
HÁBITOS CELULARES.

El cercamiento gradual del receso metabólico
llevaría al hábito celular (donde la vacuola
protónica ya no es necesaria). |
|
EL LIBRO.
 |
Haga clic aquí
para bajar ECOPOESE- A criação da
ecosfera, 2ª Ed., 2006; (en portugués)
- PDF Gratuito (1,7 Mb, 345 páginas).
O deje un mensaje
para comprar la versión impresa.
Esta obra es protegida por derecho
autoral. Sin embargo, cualquier parte del documento
se puede reproducir o traducir libremente para propósitos
no comerciales, con tal que la fuente se refiera
claramente.
Este libro debe ser referenciado
en el siguiente modo:
Félix de Sousa, Raul A., (2006). Ecopoese
- A criação da ecosfera, 2ª Ed.,
Rio de Janeiro. |
UN ARTÍCULO.
Una versión condensada viXra
en inglés puede ser bajada aqui
( PDF Gratuito 526 kb, 39 páginas ).
Para la referenciar:
Félix de Sousa, Raul A. (2010). The
Ecopoesis Model: Did Free Oxygen Fuel the Origin of Life?
viXra: 1010.0001
|
 ECOPOIESIS
ECOPOIESIS