Vôtre opinion
Autres Langues
English
Español
Interlingua
Português
|
|
ÉCOPOÈSE.
Resumé -
Le modèle de l'Écopoèse est
une hypothèse au sujet de l'origine de
vie sur la Terre qui propose que les cycles géochimiques
des éléments biogèniques
(ou cyles biogéochimiques) aient précédé
l'apparaissance des organismes. L'existence de
tels cycles dans un monde protobiologique implique
la présence de forts doneurs (réducteurs)
et accepteurs (oxydants) d'électrons, ravitaillés
par des réactions abiotiques dans l'environnement
aqueux primitif de la planète.
Le cyclage des composés de carbone, d'azote
et de soufre aurait lieu dans les océans,
un environnement aqueux en contact avec une lithosphère
riche d'ions réducteurs tel que le fer
ferreux (divalent) et le sulfure, et une atmosphère
contenant de quantités substantielles d'oxygène
moléculaire (libre), produit par la photolyse
atmosphérique de la vapeur d'eau.
Les transitions redox appartenant aux processus
vivants seraient donc des attributs primordiaux
de l'ecosphère naissante, en déterminant
la nature de matériaux et des processus
d'obtention d'énergie qui façonneraient
progressivement l'évolution de l'organismalité.
Les hauts niveaux atmosphériques de bioxide
de carbone expliqueraient quelques propriétés
réactives essentielles de ce monde protobiologique,
telles que la possibilité de fixation du
carbone, et la propagation chirale.
Le modèle de l'Écopoèse
(Félix de Sousa, R.A., 2000, 2006) pour
l'origine de la vie est basé sur l'idée
qu'une atmosphère contenant un haut pourcentage
d'oxygène moléculaire, produit par
la photolyse atmosphérique de la vapeur
de l'eau, est un trait essentiel de l'écosphère
primitive de nôtre planète.
L'écopoèse offre une perspective
différente par rapport aux autres modèles
de biopoèse. Traditionnellement, l'origine
de la vie est considérée équivalente
à l'heureuse apparition d'organismes cellulaires
très simples, dont l'évolution métabolique
conduirait dorénavant l'évolution
générale de l'environnement (l'atmosphère
riche en oxygène serait un résultat
de ce procès). Dans l'écopoèse,
en contraste, c'est l'environnement physique qui
joue le rôle principal, et non pas les organismes.
En plus de douer l'environnement, dés le
debut, avec la protection continuelle d'une couche
d'ozone, la présence précoce de
l'oxygène libre détermine l'établissement
d'un flux d'électrons, qui laissent les
constituants réducteurs de la lithosphère,
croisant les mers archéennes, pompées
par la production photolytique d'oxygène
dans l'atmosphère de la Terre. Ce flux
est intermedié et en grande partie canalisé
par les transitions redox des élements
redox-sensibles dans l'hydrosphère.
Ces interactions de grande échelle de l'environnement
causent le développement d'un métabolisme
de base géochimique dans un protoplasme
planétaire (holoplasme) tout en créant
des conditions pour l'évolution graduelle
de la vie organismale.
|
La large différence
de potentiels électrochimiques dans l'environnement
primordial causerait l'apparition des cycles géochimiques
des éléments biogéniques. Les
voies métaboliques primitives sont ainsi
provenantes de l'interaction de ces cycles et leurs
produits. La génération d'ordre dans
le système survient des transitions énergétiquement
favorables, notamment dans l'oxydation de matière
organique, et des propriétés physiques
et chimiques des composés impliqués.
Ce protométabolisme planétaire est
essentiellement congruent avec la biochimie d'aujourd'hui,
y comprises la fixation du carbone et de l'azote,
et la dégradation aérobique des composés
organiques (oxydation pleine à CO2).
L'évolution biologique, comme une règle,
poursuivrait par un contrôle utilitaire croissant
de telles réactions, plutôt que par
leur création. Un métabolisme géochimique
nu aurait donc évolué de manière
congruente envers nos procès enzymatiques
modernes.
|
CYCLES GÉOCHIMIQUES
DES ÉLÉMENTS BIOGÉNIQUES DANS LA
PÉRIODE ARCHÉENE.

|
La
rangée des potentiels électrochimiques
dans la Terre protobiologique est calculée
comme s'étendant dès les valeurs du
fer bivalent lithosphérique à ceux
de l'oxygène atmosphérique.
Les premiers cycles géochimiques sont constitués
par les espèces chimiques possibles pour
les éléments biogéniques et
par leurs transitions dans cet intervalle.
La nature oxydante de l'atmosphère primitive
serait déterminée par la production
abiotique de l'oxygène, provenant de la photodécomposition
de la vapeur d'eau atmosphérique suivie par
la perte gravitationnelle d’hydrogène.
La très basse solubilité de l'oxygène
dans les océans primordiaux chauds et le
manque de terres émergées exigeraient
la maintenance d'une concentration élevée
de l'oxygène dans l'atmosphère, pour
que le flux des électrons provenant des compartiments
hydrosphérique et lithosphérique ait
pu compenser la perte par la photodécomposition.
La plupart des gaz réducteurs dérivés
de l'activité volcanique seraient fournis
directement à l'hydrosphère ou auraient
une résidence courte dans l'atmosphère.
Un peu de méthane et de monoxyde de carbone
moins solubles, cependant, auraient pu coexister
dans un équilibre avec l'atmosphère
oxydante.
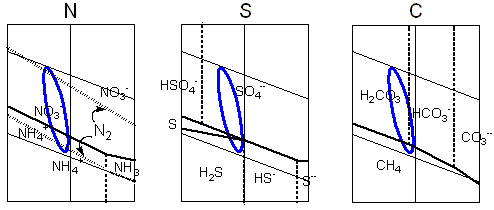
Les régions de stabilité pour les
espèces chimiques dérivées
des éléments biogéniques sont
tracées dans des diagrammes de Pourbaix.
La ligne bleue relie les valeurs extrêmes
de pH et de potentiel électrochimique calculés
pour le milieu hypercarbonique. Elle prétend
illustrer, dans une vue approximative, les transitions
chimiques possibles dans les conditions protobiologiques
proposées. Dans le cas du soufre les cycles
seraient encore compliqués par les interactions
avec le fer bivalent. Le diagramme de Pourbaix pour
le carbone n'inclut que les deux extrémités,
l'espèce la plus réduite- le méthane,
et la plus oxydée, l'anhydride carbonique
(et ses dérivés aqueux, l'acide carbonique,
l'ion bicarbonate et l'ion carbonate), à côté de
plusieurs composés possibles exhibant des
états intermédiaires de réduction. |
|
| |

|
|
Les
propriétés carboxylantes et condensantes
pretées au milieu aqueux (dit «hypercarbonique»)
par la présence d'une lourde atmosphère
de CO2 sont particulièrement
importantes, puisqu'elles permettent la fixation
du carbone dans la présence du pouvoir réducteur,
et la propagation chirale.
|
UN MONDE HYPERCARBONIQUE.
| |
L'expression
«milieu hypercarbonique» décrit
les propriétés réactives
de l'environnement océanique de la Terre
primitive sous la composition atmosphérique
postulée dans le modèle de l'écopoèse.
Grâce à la grande quantité
de CO2 dissoute, des espèces
chimiques telles que l'acide carbonique, l'ion
bicarbonate et les ions bivalents (surtout l'alcalino-terreux
magnésium) se présentent dans des
concentrations beaucoup plus hautes que dans les
océans d'aujourd’hui, prêtant
au milieu des propriétés carboxylantes
capables d'affecter les groupements amino et les
carbanions. Sous l'équilibre carboxylation-decarboxylation ainsi determiné, plusieurs composés organiques formeraient des séries d' «analogues hypercaboniques» étroitement ressemblants dont les membres ne diffèrent que quant à la quantité de CO2 ajouté à ses molecules.

Le
milieu très polaire, avec une baisse activité
d'eau (Aw) gouverne l'équilibre
hydratation-déshydratation. En plus, l'équilibre
réduction-oxydation fait partie de l'ensemble
de l'environnement océanique, placé
entre une lithosphère qui fournit des ions
réducteurs (surtout le fer et le soufre bivalents)
et l'atmosphère oxygénique. Les facteurs
qui déterminent l'équilibre de carboxylation-décarboxylation
et l'équilibre hydratation-déshydratation
sont nommés des facteurs permanents et sont
liés à des conditions qui ne flottent
que très légèrement. L'équilibre
réduction-oxydation, par contre, est dirigé
par des facteurs éventuels,qui varient fortement
entre les extrémités redox de l'environnement
protobiologique.
La chimie du milieu hypercarbonique est centrale
pour le modèle de l'écopoèse
puisqu'elle concilie, tel qu'il est suggéré
par le principe de la congruence, la relevance biologique
des réactions de carboxylation et de décarboxylation,
et les évidences géologiques d'une
paléoatmosphère lourde de CO2. |
|
Lorsque
le dioxyde de carbone est exceptionellement stable
vis-à-vis la réaction de réduction
dans les solutions aqueuses, la fixation du carbone
dépendrait de la carboxylation des composés
de carbone préexistants. Les cycles protobiologiques
du carbone auraient, dans le sens anabolique, des
réactions de carboxylation intercalés
avec des étapes de réduction. Inversement,
les cycles cataboliques alterneraient des décarboxylations
et des étapes d'oxydation, tel que dans les
cycles métaboliques modernes. Les possibles
analogues non-enzymatiques des ces réactions
sont discutés avec un certain détail
dans le livre, aussi que les mécanismes par
lesquels l'équilibre réduction-oxydation
dans l'écosphère protobiologique serait
fermement lié aux processus de carboxylation
et décarboxylation.
|

La fixation du
carbone exigerait au moins un modeste apport (dit
«primaire») d'hydrocarbures dérivés
des carbures lithosphériques, notamment l'acétylène,
qui pourrait être converti directement, à
travers l'hydratation et la carboxylation, aux acides
pyruvique et oxaloacétique, composants essentiels
du noyau central des voies biochimiques. La participation
de ces corps suggère le cycle de Krebs bidirectionnel
comme le trait principal de la circulation de carbone
dans l'écosphere primordiale.
La disponibilité du pouvoir réducteur
pour les processus anaboliques (dérivé
principalement du fer divalent et des sulfures lithosphériques),
comparée à celle de l'oxygène
atmosphérique à peine soluble, favoriserait
l'accumulation des composés organiques dans
l'hydrosphère.
Le questionnement au sujet de la nature de ces composés
et de ses processus de formation peut être
aidée par la présomption d'une évolution
congruente.
|
LE PRINCIPE DE LA
CONGRUENCE.
| |
Le principe
de la congruence est un outil heuristique censé
pour guider la recherche de l'histoire des systèmes
vivants. Il énonce simplement que la vie
moderne est une référence utile et
nécessaire dans l'étude des étapes
précédentes de la vie. La congruence
peut être envisagée comme complément
à la théorie évolutionnaire
puisqu'elle représente la recherche des évidences
d’ancestralité commune à un
niveau biochimique.
Bien que les similitudes biochimiques parmi les
organismes existants soient généralement considerés une preuve non ambiguë
de leur évolution commune, les modèles
classiques pour l'origine de la vie proposent un
monde précédent, «prébiotique»,
où les composés organiques se seraient
accumulés par des processus étranges
à la biochimie d'aujourd'hui.
Les premières propositions théoriques
concernant la recherche de la nature évolutionnaire
du métabolisme ont été présentées
par Granick dans les années 40, et ont été
appliquées plus tard, notamment par De Duve
et par Morowitz, au champ de l'origine de la vie.
Le modèle de l'Ecopoèse prolonge davantage
le domaine de la congruence, de l'évolution
des organismes à celle de l'écosphère
dans l'ensemble.
Cette conception implique que les cycles géochimiques
des éléments biogéniques ont
toujours été essentiellement identiques,
mais également que les raides gradients d'énergie
qui nourrissent les processus vitaux n'ont pas été
créés par les organismes. Au contraire,
c'est l'existence même des processus générateurs
d'énergie dans l'environnement primordial,
particulièrement la dégradation aérobique
de la matière organique (oxydation totale
à CO2), qui aurait
été essentielle à la création
de l'ordre inhérente à la vie organismale.
L'utilization du terme «protobiologique»
impliquant la vie primitive, au lieu de «prébiotique»
(avant la vie) emphatise le type de continuité
qui caractérise le principe de la congruence.
Elle est cohérentment suivie dans le modèle
de l'écopoèse. |
|
La congruence exige
l'analogie chimique entre ces deux étapes
de l'évolution biochimique, et le parallelisme
entre l'action des facteurs protobiologiques de
l'environnement et celle des coenzymes est mis en
évidence.
|
PROCESSUS CONGRUENTS
DU MONDE HYPERCARBONIQUE.
| |
Facteurs réactifs de l'environnement et réactifs métaboliques correspondents.
1 - Permanents.
Facteur de l'environnement (et réaction contraire) |
Réactifs métaboliques correspondents |
Origine |
+ H2O
- H2O
|
(spontané)
ATP, GTP, etc. |
Hydroatmosphère
|
+ CO2
- CO2
|
Biotine
(spontané) |
Hydroatmosphère
|
2 -Éventuels primaires.
[H]
[O]
|
TH - Transporteurs Réduits
(NADH, FADH2, etc.)
T - Transporteurs Oxydés
(NAD+, FAD, etc.) |
Pouvoir Reducteur Lithosphérique
Oxygène Atmosphérique |
+
H2S
- H2S
(+H2O)
|
CoenzymeA-SH
(spontané) |
Hydroatmosphère (Cycle du S) |
+
NH3 (NH2COOH)
- NH3
(+H2O)
|
Composés aminés
(spontané) |
Hydroatmosphère (Cycle du N) |
+
H2PO4-
- H2PO4-
(+H2O) |
ATP, GTP, etc.
(spontané) |
Hydroatmosphère |
3 - Éventuels secondaires.
H2S2 |
Acide Lipoïque |
H2S
/ [O] |
NH2COOH
/ [H]
(NH2COSH / [H])
Nucléophile d'Azote |
Pyridoxamine |
NH2COOH
/ [H] |
CN-
Nucléophile de Carbone |
Thiamine |
Cyanure
ou derivés de l'acide formique (Un-carbone) |
|
|
Les coenzymes représentent un premier
pas envers l'incorporation du métabolisme
nu primordial dans des unités portant de
l'activité catalytique. La probabilité
que cette transition se soit passée surtout
pendant le "monde de l'ARN", où
les fonctions métaboliques étaient
réalisées par des ribozymes, est
documentée par le fait que ces coenzymes
sont, pour la plupart, structurellement relationnés
avec les nucléosides contenant du ribose.
La propagation chirale (polymérisation
enantioselective) assurerait la synthèse
de polypeptides homochiraux à travers la
formation de centres prochiraux dans les dérivés
hypercarboniques des matériaux peptidiques.
|
PROPAGATION CHIRALE DANS
LE MILIEU HYPERCARBONIQUE.
| |

Le
mécanisme de propagation chirale ci présenté
pour les aminoacides racémiques est
une version minimalement dénudé des
formes d'équilibre possibles dans le milieu
hypercarbonique. Conformément, la N-carboxylation
et la protonation du carboxyle de la chaîne
principale sont essentielles, mais les vraies espèces
réactives peuvent demander l'addition de
CO2 ou de H2S
au matériel peptidique, avec l'usage des
aminodiacides ou thioacides correspondants. Hormis
le fait que les organismes modernes utilisent des
aminoacides optiquement actifs, ce mécanisme
est autrement tout à fait congruent avec
la biochimie actuelle.
Dans le milieu hypercarbonique les aminoacides sont
dépourvus de propriétés zwitterioniques.
L'attaque nucléophilique du groupement amino
carboxylé au carboxyle protoné procède
facilement, donnant origine a un adduit qui contient
un carbanion stabilisé par ressonance. La
recombinaison asymétrique avec le proton
est orientée par l'aminoacide ajouté
dans le cycle antérieur. Puisque l'initiation
peut être faite par l'acylation avec un acide
carboxylique achiral, le premier aminoacide de la
chaîne pourra être n'importe quel des
deux énantiomères, mais il ira déterminer
l'orientation chirale de tous ses subséquents,
donant lieu à la synthèse de polypeptides
homochiraux.
Un autre aspect important c'est que l'acylation
du groupement amino du fragment d'aminoacide cause
l'élévation du pka du groupement carboxyle,
ou la hausse de la probabilité qu'il soit
protoné, favorisant ainsi l'allongement des
chaînes déjà en cours de formation,
plutôt que l'attaque aux monomères.
La théorie mécanistique de l'origine
de la chiralité dans le monde biologique
est un partie integrante du modèle de l'écopoèse. |
|
Les caractéristiques
organismales de la vie sont acquises d'une manière
lente et progressive et les cellules sont des structures
assez tardives dans ce modèle.
|
ÉVOLUTION
DE L'ORGANISMALITÉ: LES HABITS PROTOBIOLOGIQUES.
| |
MATERIELS PROTOBIOLOGIQUES
ET L'HABIT FLOCULAIRE.

L'agrégation initiale de matière organique
(appelée «habit floculaire»),
contrariant la tendence entropique,
est attribuée à une combinaison de
facteurs:
1) L'activité
de l'eau baissée (Aw)
du milieu hypercarbonique.
2) La coalescence
des composés hydrophobes (minimisant l'énergie
due à la séparation des charges dans
le milieu très polaire).
3) La cinétique
de formation des thioacides et leur extraction partielle
dans la phase lipide.
4) La stabilisation
de la micelle hydrophobique à travers la
formation de composés amphiphiliques à
sa surface.
5) La formation
facile des chaînes peptidiques due à
la destruction du caractère zwitterionique
des acides aminés à travers leur N-carboxylation.
6) L'adhésion
de la fraction peptidiques aux composés amphiphiliques
de la surface des micelles due à l'affinité
de charges, avec la possible participation de cations
polyvalents.
7) L'adsorption
des composés organiques de bas poids moléculaire
à la fraction peptidique.
Toutes ces
propriétés auraient contribué
à stabiliser les agrégats colloïdaux
dans la solution diluée. Elles sont même
compatibles avec la vision qui propose la Terre
primitive comme un «monde d'eaux», dépourvu
de surfaces d'évaporation considérables.
La solubilité préférentielle
de l'oxygène dans la phase lipide favorise
les processus d'oxydation auprès de la surface
de la micelle. Un processus plutôt simple,
la phosphorylation homogène - analogue à
la phosphorylation au niveau du substrat, exigeant
l'oxydation du sulfure d'hydrogène à
dissulfure d'hydrogène (analogues de l'acide
lipoïque), causerait l'oxydation des a-cétoacides
à thioacides, permettant la synthèse
de composés de «haute énergie».
Les aminoacides contenant du soufre dans la fraction
peptidique pourraient se lier au fer divalent d'origine
lithosphérique. Ces groupements fer-soufre,
ancêtres des ferredoxines d'aujourd'hui, sont
les responsables pour la localisation du pouvoir
réducteur, concentrant des potentiels redox
dans les agrégats colloïdaux primitifs,
qui auraient ainsi des pôles réducteurs
et oxydants.

Une interaction initialement souple et générique
entre les polypeptides et les polymères phosphodiester
(PPDE - ou ARN primitif) augmenterait en spécificité
donnant lieu à la traduction, et au contrôle
structurel et utilitaire qui caractérise
l'«habit réticulaire».
L'HABIT RÉTICULAIRE.

Bien que la phosphorylation
hétérogène («respiration»)
exige des barrières physiques, telles que
les membranes, elle pourrait être développée
avant l'avènement de l'«habit cellulaire»,
lorsque les protons qui résultent de l'oxydation
des transporteurs d'électrons solubles en
lipides auraient étés ségrégués
dans des vacuoles protoniques (ou acides), créant
ainsi un potentiel entre cette vacuole et le compartiment
extérieur (ou métabolique), ce qui
fournirait un moyen plus effectif pour la synthèse
des composés de haute énergie.
CELLULARITÉ
ET RESPIRATION CELLULAIRE.

Dans le modèle
de l'Écopoèse la cellularité
n'apparait pas comme une méthode pour protéger
les composés organiques de la dilution, mais
pour promouvoir leur oxydation et pour en profiter.
Cette proposition d'une aurore aérobique
pour la cellularité est fondamentée
sur les propriétés fonctionnelles
énergétiques de la cellule plutôt
que sur son apparence physique.
HABITS CELLULAIRES.

La
clôture graduelle du recès métabolique
mènerait à l'habit cellulaire (où
la vacuole protonique n'est plus nécessaire). |
|
LE LIVRE.
 |
Cliquez ici
pour télécharger ECOPOESE - A criação
da ecosfera, 2ª Ed., 2006; (en portugais) -
PDF Gratuit (1,7 Mb, 345 pages).
Ou laissez un message
pour acheter la version imprimée.
Ce document est protégé
par des droits d'auteur. Cependant, des parties
quelconques de l'œuvre peuvent être librement
reproduites ou traduites pour des buts non-commerciaux,
pourvu que la source soit clairement référencée.
Ce livre doit être référencé
de la manière suivante:
Félix de Sousa, Raul A., (2006). Ecopoese
- A criação da ecosfera, 2ª Ed.,
Rio de Janeiro. |
UN ARTICLE.
Une version abregé viXra
en anglais peut être télécharchée
ici ( PDF Gratuit
526 kb, 39 pages ).
Pour la référencer:
Félix de Sousa, Raul A. (2010). The
Ecopoesis Model: Did Free Oxygen Fuel the Origin of Life?
viXra: 1010.0001
|
 ÉCOPOÈSE
ÉCOPOÈSE
